ヒトDNA組換えタンパク質ダブルリング構造を解明- 配偶子形成の最も重要な相同組換え反応はリング内で -(プレスリリース)
- 公開日
- 2004年05月07日
- BL44B2(理研 物質科学)
平成16年5月7日
独立行政法人理化学研究所
|
独立行政法人理化学研究所(野依良治理事長)は、親から子へ遺伝子が伝わる過程で中心的な役割を果たす相同DNA組換え反応※1を行うヒトのタンパク質「Dmc1」の立体構造を世界で初めて決定し、相同DNA組換えの分子メカニズムを解明することに成功しました。理研ゲノム科学総合研究センター(榊佳之センター長)タンパク質構造・機能研究グループの横山茂之プロジェクトディレクター、杵渕隆研究員、胡桃坂仁志客員研究員(早稲田大学理工学部助教授)らの研究グループによる成果です。 (論文) |
1.背 景
減数分裂は、真核生物※2が子孫を残すために生殖細胞で行う特殊な細胞分裂です。この分裂が起きるときに、DNA組換え反応が起きて、両親から受け継いだ染色体(相同染色体)が対合し、同じ染色体部位に位置する遺伝子間で相同的対合反応(図2)が行われます。この相同的対合反応を経由した一連の反応を相同DNA組換え※1といいます。生物はこの反応を用いて、両親から受け継いだ遺伝情報を交換し、新しい組み合わせの遺伝子セットを持った子孫を作って遺伝的多様性を確保しています。また、この相同DNA組換え反応は、傷ついたDNAの修復にも使われており、細胞が正常に機能するために不可欠なメカニズムです。
研究グループでは、減数分裂期での相同的対合反応を行うDmc1に着目し、その全長タンパク質の立体構造を決定することに成功しました。そして解明された立体構造に基づいた生化学的解析によってDmc1による相同DNA組換え作用機構の解明を試みました。
2.研究の手法
Dmc1は、341アミノ酸からなり、単鎖DNA、二本鎖DNAの両方に結合し、相同的対合反応を促しますが、その立体構造、分子反応メカニズムは明らかになっていませんでした。研究グループは、まず大腸菌体内でDmc1を生産し、様々な精製技術を使い、不純物のない状態までDmc1を精製しました。そして、精製されたDmc1を用いて単結晶(約600μm)を作製し(図3)、大型放射光施設(SPring-8)の理研構造生物学 II ビームライン(BL44B2)を用いてその立体構造を原子レベルで決定しました。
3.立体構造の解析結果から明らかになったこと
SPring-8からの高輝度なシンクロトロン放射光により、最終的には3.2ÅのX線回折データを得ることができました。これらのデータよりDmc1の立体構造を決定し、Dmc1が巨大なリング構造を形成することが明らかになりました。さらに、立体構造に立脚した生化学的解析により、そのDNA結合領域を同定することに成功しました。
- 1)「Dmc1は、16個のユニットからなる巨大なダブルリング構造を形成している」
Dmc1はバクテリアの組換えタンパク質であるRecAのヒトホモログ※3です。これまでに、バクテリアのRecA(図4)はらせん状のフィラメント構造を形成して相同DNA組換えを行うことが知られていました。研究グループが解明したDmc1の分子構造から、Dmc1は、ホモログであるRecAとは異なり、8個のユニットからなるリング構造が重なった16ユニットからなるダブルリング構造を形成して機能することが明らかになりました。ダブルリングの大きさは、縦が約85Å、横が約130Åです。このような巨大なダブルリング構造の報告は、今までに例がなく、今回のケースが世界で最初の報告となります(図5)。
- 2)「Dmc1は、なぜ安定なリング構造を形成するか?」
Dmc1には、双子のようにアミノ酸配列が非常に似ているRad51(図4)というタンパク質が存在します。ところが、不思議なことにRad51は、Dmc1とは異なり、RecAときわめてよく似たらせん状のフィラメント構造を形成します。なぜこの双子タンパク質が、リングとらせん状フィラメントといった異なった高次構造を形成するのか?という謎が、今回の研究によって明らかになりました。Dmc1は、リング構造中でユニット間に3本の水素結合を形成していますが、これらの水素結合を形成するために必須のアミノ酸がRad51には存在しません。そのため、Dmc1は3本の水素結合によって安定なリング構造の形成が可能になり(図6)、一方、Rad51はDmc1と類似のリング構造を形成するのに適していないことが分かりました。
- 3)「相同DNA組換えの反応中間体モデル」
Dmc1のダブルリング構造には、中心に直径30Åの穴が、ダブルリング構造の側面に直径15Åの穴が開いていることが分かりました。中心の直径30Åの穴は、二重鎖DNAを収容するためにちょうど良いサイズで、側面の直径15Åの穴は単鎖DNAを収容するためにちょうど良いサイズであることが分かりました。これらの穴にDNAが直接結合することを、変異体Dmc1を用いた生化学的解析により明らかにしました。これらの結果から、単鎖DNA-二重鎖DNA-Dmc1からなる三者複合体モデルを提案しました。(図1)。このモデルでは、中心の穴に二重鎖DNAが結合し、リング側面の穴に単鎖DNAが結合します。そして、ダブルリング構造の内部で相同対合反応が行われ、中心の穴から組換えが終了したDNAが出てきます。これらのモデルは、らせん状フィラメント構造によって組換えを行う従来のRecAタイプとは全く異なった、新しいDNA組換え反応機構を示しています。従って,減数分裂期の相同DNA組換え反応には,RecA・Rad51タイプとDmc1タイプの2種類があることがわかりました.このように生物は,複数の相同DNA組換え機構を持つことにより効率良く多様性を獲得してきたと考えられます。
4.今後の展開
本成果によって、今まで不明であった減数分裂期の相同DNA組換え分子機構の核心部分が明らかになりました。現在、進化によって地球上に多種多様な生物が生存し、繁栄しているのは、相同DNA組換え反応の結果であり、従って、本研究成果が基礎となって、細胞内でのDNA組換え効率を人為的に増進させる方法が開発される可能性があります。そのことは、今後、染色体上での遺伝子治療の基礎技術や、農作物や家畜などの品種改良をはじめとする、医療や産業の分野に大きく貢献することが期待されます。
<参考資料>
Dmc1ダブルリング(黄色)の中心の穴から向かって、上から二重鎖DNA(赤色)が通っている右のリング-リング間の隙間の穴から単鎖DNA(青色)が入ってきて、リングの中心で相同性対合反応が起こり、下の穴から組換え反応が終わった二重鎖DNAが出てくる。さらに組換わらなかった二重鎖DNAの反応側の鎖は、左のリング間の隙間からでる。
2つのタンパク質は、Dmc1とは対照的ならせん状の構造をとる。このモデルは、電子顕微鏡の解析から明らかにされた。
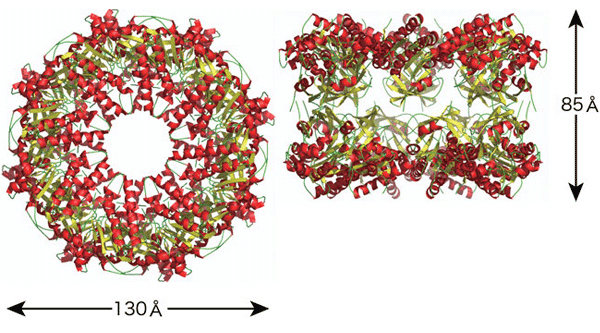
上から見た図(左)と横から見た図(右)
左の図は、8個のユニットからなるDmc1のリング構造、各ユニットはそれぞれ別の色で示した。
右の図は、拡大した3本の水素結合である。水素結合は、ピンク色で示した。この水素結合が各ユニット間に存在し、リング構造を安定化している。
<補足説明>
- ※1 相同DNA組換え
2分子のDNAの間でおこる組換え反応の内、塩基配列の相同性に依存して行われるものを指す。生体内では、精子や卵子などの配偶子を形成する過程で必須であり、両親のゲノムDNAの情報の再編成の過程で行われ、さらに、DNAの二重鎖切断を修復する。
- ※2 真核生物
生物の分類のひとつであり、真核細胞からなる生物を指す。真核細胞の最も際立った特徴は、細胞内に細胞核を持ち、細胞のそれ以外の部分からは膜で区切られていることである。核の中には遺伝情報を保持しているDNAが収められている。
- ※3 ホモログ
2つのタンパク質を比較すると、立体構造(フォールド)は似ているにも関わらず、アミノ酸の一致度は低い場合が多く存在する。これらのタンパク質ペアは遠縁のホモログと呼ばれ、弱い進化的類縁関係を有する。
|
<本件に関する問い合わせ先> 研究員 杵渕 隆 客員研究員 独立行政法人理化学研究所 (報道担当) <SPring-8についての問い合わせ先> |
- 現在の記事
- ヒトDNA組換えタンパク質ダブルリング構造を解明- 配偶子形成の最も重要な相同組換え反応はリング内で -(プレスリリース)