細菌べん毛のミクロのプロペラが形態をスイッチするナノ機構を解明(プレスリリース)
- 公開日
- 2010年03月15日
- 電子顕微鏡(大阪大学)
2010年3月15日
大阪大学
理化学研究所
|
国立大学法人大阪大学(鷲田清一総長)と独立行政法人理化学研究所(野依良治理事長)は、下記の共同研究チームが、低温電子顕微鏡法とらせん像再構成法により細菌の遊泳器官であるべん毛の超分子繊維構造を解析し、微小な生体プロペラの形成とスイッチの分子メカニズムの解明に成功したことを発表します。これは、理研放射光科学総合研究センター(石川哲也センター長)タンパク質結晶構造解析研究グループの眞木さおり研究員、米倉生体機構研究室の米倉功治准主任研究員、大阪大学生命機能研究科の難波啓一教授の共同研究による成果で、米国科学誌「Nature Structural & Molecular Biology」の電子版で2010年3月14日(英国ロンドン時間)に公開されます。 (論文) |
《研究の背景》
細菌の運動器官であるべん毛は、約30種類の異なったタンパク質から構成されるナノマシンで、各部の構造は人工の電気モーターに驚くほど良く似ていています。回転子、固定子、反転制御装置、軸受け、自在継ぎ手、プロペラに相当する構造を持ち、これらを構成するタンパク質「パーツ」は、自ら組み上がる自己組織化能力を持っています(図1)。べん毛繊維は一種類の蛋白質フラジェリンが2万 ~ 3万個もらせん状に重合して形成するミクロのプロペラで、その長さは菌体長の約10倍、10 ~ 15μmにも達します。細菌の遊泳時には、べん毛繊維は緩やかな曲率を持つ超らせんと呼ばれる形態をとっていますが、餌となる化学物質に向かったり水温の低い場所から逃げたりする際は、1分間に約2万回もの高速回転をしているモーターが1ミリ秒程度で逆転し、この時、べん毛繊維のミクロの左巻き超らせん構造は一時的に右巻きに変換します。この形態変換機構には、L型とR型とよばれる、フラジェリン分子の2つの異なる立体構造が鍵を握っていることが知られていましたが、その詳細なメカニズムは謎のままでした。
《研究内容と成果》
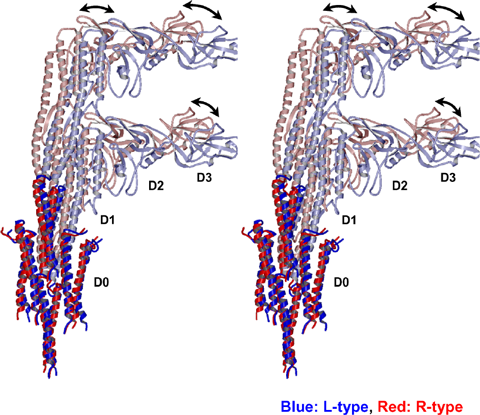
今回の研究成果を発表したグループは、2003年にR型の繊維構造を報告しましたが、その後も低温電子顕微鏡法とらせん像再構成の技術開発を進め、今回はL型繊維の原子モデルの構築に成功しました(図2)。その結果、L型とR型の構造の違いによって引き起こされるべん毛繊維の形態変換機構に関して、その分子メカニズムを明らかにしました。らせん繊維の形態変換を実現していたのは、べん毛繊維の中心にある二重円筒構造の内側の緊密な分子間相互作用と外側の柔軟な分子間相互作用で、生物ナノマシンの持つ柔らかい制御の仕組みが明らかになりました(図3)。こうして1960年代から謎であった、細菌の遊泳をコントロールするしくみの一端を解明することができました。今回の成果は、将来のナノテクノロジーの応用として、ナノモーターのプロペラの設計などにも役立てることができると期待されます。
《今後の展開》
繊維構造を形成する超分子複合体は生体内に数多くありますが、結晶の作製が困難で、X線結晶解析法による高分解能での分子構造解析は極めて困難です。今回の結果は、結晶を作成することなく、生体超分子の自然な機能構造を解析できる手法の開発という点でも、きわめて重要であると考えられます。
《参考資料》

サルモネラ菌や大腸菌は数本のべん毛を細胞周辺から延ばし、束にして回転させて泳ぐ。べん毛はらせん型のスクリュープロペラで、らせんのピッチは2.5 μm、直径は0.5 μm。それぞれの根元に回転モーターを持つ。べん毛は大まかに回転モーターである基部体、ユニバーサルジョイントであるフック、プロペラのように動くべん毛繊維の、3つの部分構造で構成される(CM;細胞膜/PG; ペプチドグリカン層/OM;外膜)。

(問い合わせ先) 米倉 功治(よねくら こうじ) 理化学研究所 播磨研究推進部 企画課 (SPring-8に関すること) |


- 現在の記事
- 細菌べん毛のミクロのプロペラが形態をスイッチするナノ機構を解明(プレスリリース)